Les neurones de la peur
Publié par Antoine Depaulis, le 22 juillet 2016 18k

La peur est une question de survie pour les espèces animales, y compris la nôtre. Face à un danger, elle permet de déclencher différentes réponses comportementales et physiologiques qui doivent être rapides et surtout adaptées aux conditions de l'environnement. Ces réactions vont de l'immobilisation à la défense active en passant par la fuite et mettent en jeu des circuits nerveux différents mais qui interagissent entre eux afin de permettre à l'individu de s'adapter à l'évolution de la situation. Un réseau de chercheurs suisses, français, portugais et américains vient d'en préciser l'architecture et de faire un pas de plus dans la compréhension des comportements d'adaptation, ou de mal-adaptation, à la peur et à sa petite soeur l'anxiété.
La peur est un sujet de recherche en Neuroscience depuis de nombreuses années. La perception d'une situation dangereuse est en effet vitale pour la survie de l'individu ainsi que la coordination motrice et les modifications physiologiques nécessaires pour s'y adapter et/ou réduire le danger. Aussi, les stratégies de défense d'un individu sont généralement assez "simples" : il peut s'immobiliser et se soustraire ainsi à la détection d'un prédateur. Se faire "oublier" en quelque sorte ! Solution simple, peu coûteuse sur le plan énergétique mais tout de même assez risquée... Il peut fuir aussi et augmenter ainsi la distance entre lui et la source de danger, stratégie très efficace mais coûteuse en énergie et qui n'est pas dénuée d'un certain risque. Enfin, la défense active est une stratégie hautement risquée et très coûteuse, mais qui permet parfois d'éliminer la source de danger et donc la peur qu'elle engendre...
La décision d'adopter l'une ou l'autre de ces 3 stratégies de défense doit être prise rapidement, presque de façon réflexe, tout en permettant d'activer, si nécessaire et tout aussi rapidement un plan "B" qui fasse appel à une autre stratégie de défense. Le choix va dépendre bien sûr de la nature du danger, de la configuration de l'environnement mais aussi de l'expérience de l'individu. Si je me trouve face à un homme armé alors que j'allais tranquillement aux toilettes dans le train, ma stratégie de défense va être différente selon que je suis un ancien marines de l'armée américaine ou un vieil homme souffrant d'arthrose...
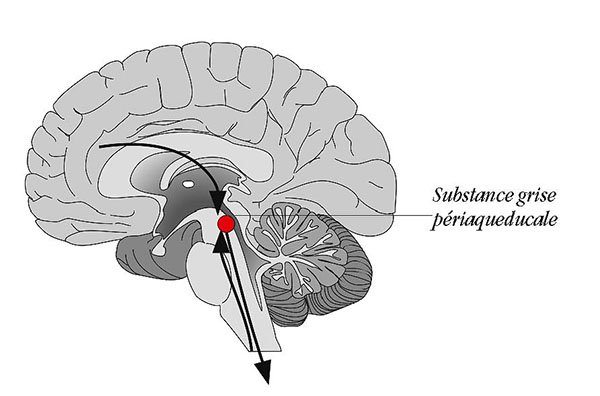
Dans les années '90, avec Richard Bandler, alors Professeur à l'Université de Sydney et grand spécialiste des réactions de défense, nous avions montré chez le rat (qui utilise avec brio chacune des trois stratégies) qu'une structure nerveuse appelée la "substance grise périaqueducale" (periaqueductal gray matter in English ou PAG pour les intimes!) possède des neurones dont la stimulation déclenche, selon leur localisation, l'une ou l'autre de ces trois stratégies. Plus précisément, et avec Pascal Carrive (actuellement Associate Professor à l'Université de Sydney), nous avions proposé que la partie latérale de cette belle substance grise - sorte de manchon de neurones autour de l'aqueduc de Sylvius, dans le mésencéphale - était organisée en colonnes orientées de façon antéro-postérieure avec des connexions différentes et des implications distinctes dans le contrôle des comportements de défense (1).
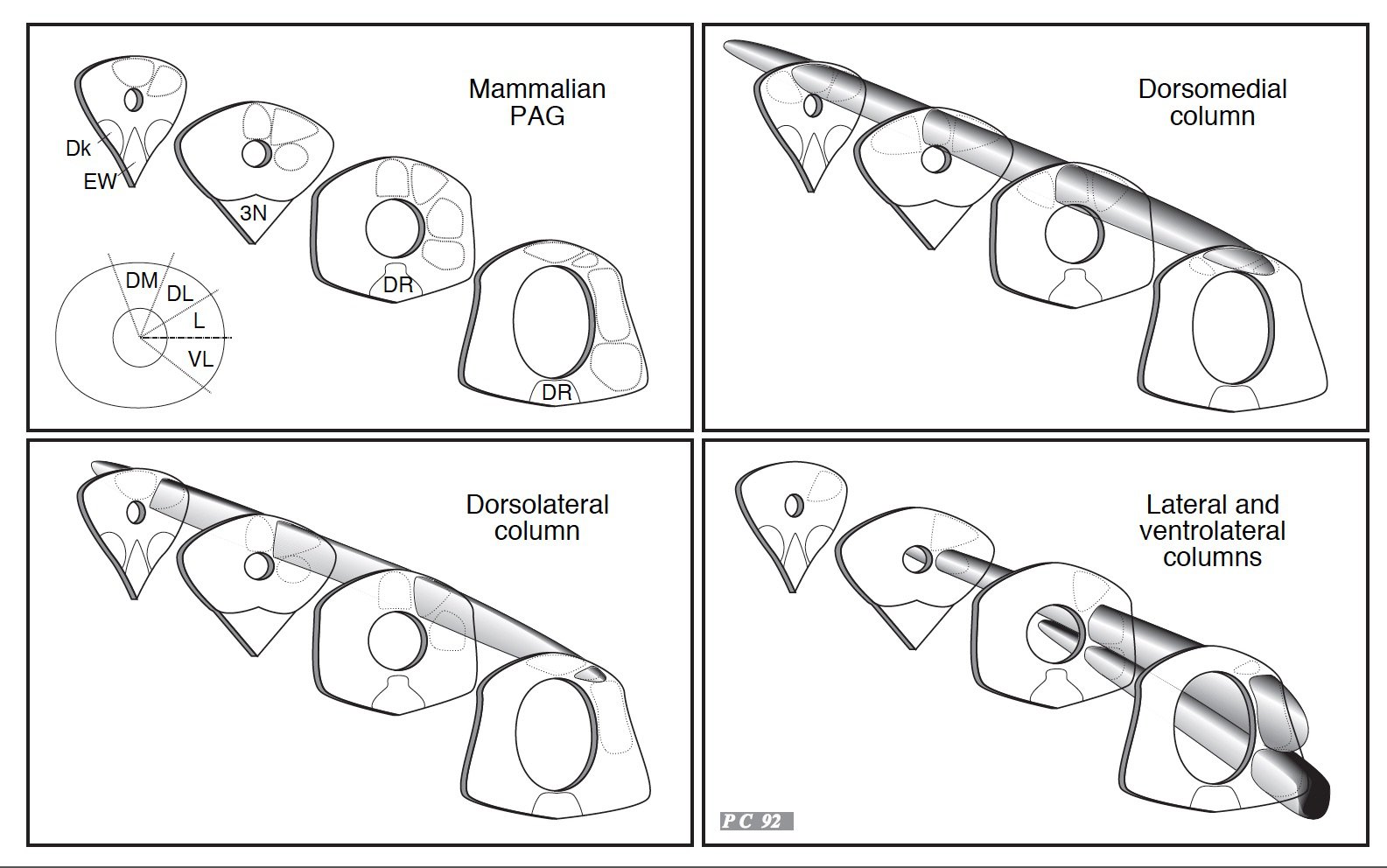
De façon schématique, la colonne la plus dorsale permettrait la coordination d'une réaction de défense, la plus ventrale celle d'une réaction d'immobilité et la colonne intermédiaire celle d'une fuite. Avec les moyens de l'époque (activation pharmacologique localisée dont la spécificité et la localisation restaient très incertaines ou marquage de l'expression de protéines révélatrices d'une activation neuronale soutenue), nous avions montré que la colonne ventrale pouvait être activée naturellement par un pincement de la nuque et induire une immobilisation (2). Par ailleurs, Richard Bandler et ses collaborateurs, avaient collecté plusieurs données suggérant que cette organisation en colonne existait également chez le chat. Ces données obtenues dans deux espèces différentes laissent penser que tous les mammifères, y compris l'homme, posséderaient une organisation semblable de leur substance grise périaqueducale. Hypothèse toutefois difficile à examiner car cette structure, très enfouie dans notre masse cérébrale est difficilement accessible aux techniques d'imagerie chez l'homme. De plus, à l'époque, l'étude de comportements aussi "basiques" n'était peut-être pas l'une des priorités dans la connaissance du cerveau.
Les choses ont changé et plusieurs travaux chez l'animal et chez l'homme ont montré au cours de ces dernières années, le rôle de certaines structures limbiques (en particulier le noyau central de l'amygdale) dans la peur et l'anxiété, avec un retour de l'intérêt pour la substance grise périaqueducale. Tout récemment, Andréas Lüthi et ses collaborateurs de Bâle, Bordeaux, Lisbonne, Standford et Harvard viennent de publier dans le journal "Nature" une étude qui révèle l'organisation des circuits nerveux qui, au sein de cette chère substance grise, coordonnent les réactions de défense et en particulier la réaction d'immobilité ou "freezing" (3).
Pour cela, ils ont utilisé des souris dont les neurones de la région ventrolatérale de la substance grise périaqueducale ont été transfectés avec un gène leur permettant d'exprimer des canaux ioniques sensibles à la lumière. La stimulation, via une petite fibre optique, par une longueur d'onde précise de ces neurones (optogénétique) induit une réaction d'immobilité chez les souris, en l'absence de toute menace. Ils ont ensuite montré avec cette technique que ces neurones sont inhibés par de petits neurones GABAergiques locaux, eux-mêmes sous le contrôle d'une voie GABAergique elle aussi, qui provient du noyau central de l'amygdale. Tiens, tiens ! On retrouve là un autre exemple de double inhibition qui se traduit par une désinhibition des neurones de la colonne ventrolatérale de la substance grise périaqueducale.
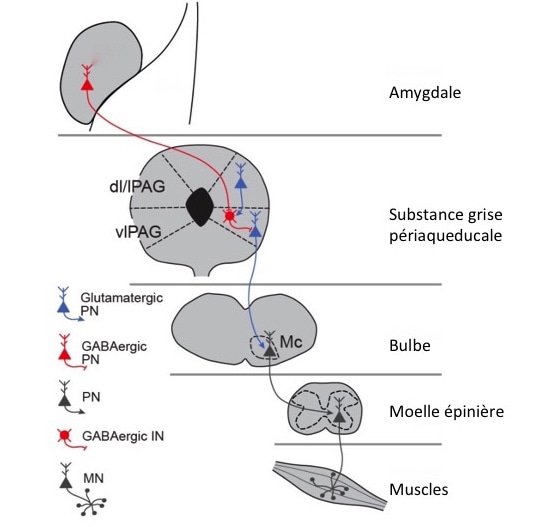
C'est bien excitant tout cette histoire de GABA! Ainsi, dans cette colonne, des neurones qui coordonnent une réaction d'immobilité pourraient être activés, pardon... désinhibés, à distance par des circuits de l'amygdale qui s'activent lors d'un danger. Grâce à une technique nouvelle qui utilise le virus de la rage "modifié" pour marquer les neurones en amont (le virus de la rage a en effet une fâcheuse tendance à "remonter" les synapses - un peu comme les saumons remontent les rivières mais pas pour les mêmes raisons - et des chercheurs malins ont exploité cette propriété pour comprendre l'organisation de circuits neuronaux (4)), l'équipe internationale a pu ainsi démontrer que les interneurones GABAergiques locaux reçoivent non seulement une projection inhibitrice de l'amygdale, mais également une projection glutamatergique (donc excitatrice) de la colonne latérale. Les neurones de cette colonne latérale sont eux impliqués dans le déclenchement d'une réaction de fuite, comportement qu'ils ont vérifié par activation optogénétique là encore. Ce circuit permettrait ainsi d'activer les neurones GABAergiques locaux et de bloquer une réaction d'immobilité qui pourrait entraver la fuite de l'animal. On voit encore que grâce au GABA tout est sous contrôle...
Cette démonstration très élégante d'un circuit neuronal impliqué dans la mise en jeu d'une réaction de défense permet de
mieux comprendre comment les circuits nerveux impliqués dans les réactions émotionnelles
peuvent déclencher une réaction de défense particulière et surtout comment des
circuits locaux permettent une réponse adaptée et rapide en face d'un danger, un peu comme un réflexe vital! On peut aussi im aginer que ces circuits s'activent parfois en l'absence de
danger soit parce qu'ils reçoivent une information erronée, soit parce que leur
sensibilité est accrue. Un comportement de défense mal adapté peut ainsi
survenir, une réaction souvent présente dans certaines pathologies
psychiatriques...
aginer que ces circuits s'activent parfois en l'absence de
danger soit parce qu'ils reçoivent une information erronée, soit parce que leur
sensibilité est accrue. Un comportement de défense mal adapté peut ainsi
survenir, une réaction souvent présente dans certaines pathologies
psychiatriques...
>>Notes
Article préparé avec la complicité de Pascal Carrive (Figure 2) et l'autorisation de Philip Tovote pour la figure 3.
- Depaulis, A., Bandler, R., 1991. The midbrain periaqueductal gray matter. Functional, anatomical and neurochemical organization, Plenum Press. ed. NATO ASI Series, New York and London.
- Keay, K.A., Clement, C.I., Owler, B., Depaulis, A., Bandler, R., 1994. Convergence of deep somatic and visceral nociceptive information onto a discrete ventrolateral midbrain periaqueductal gray region. Neuroscience 61, 727–732.
- Tovote, P., Esposito, M.S., Botta, P., Chaudun, F., Fadok, J.P., Markovic, M., Wolff, S.B.E., Ramakrishnan, C., Fenno, L., Deisseroth, K., Herry, C., Arber, S., Lüthi, A., 2016. Midbrain circuits for defensive behaviour. Nature 534, 206–212.
- Bergami, M., Masserdotti, G., Temprana, S.G., Motori, E., Eriksson, T.M., Göbel, J., Yang, S.M., Conzelmann, K.-K., Schinder, A.F., Götz, M., Berninger, B., 2015. A critical period for experience-dependent remodeling of adult-born neuron connectivity. Neuron 85, 710–717.

